Introduction
Infertility is a significant global health problem, with approximately 50% of infertility cases due to male factors. Numerous studies have identified oxidative stress and apoptosis as critical mechanisms underlying male infertility [1]. Primary somatic cells of the testes are the chief sites of androgen biosynthesis, playing a significant role in the development of male reproductive organs and spermatogenesis. Moreover, oxidative stress and apoptotic pathways are closely linked to testicular function [2].
Among tobacco components, nicotine is the most potent addictive and toxic compound, contributing to the pathogenesis of cardiovascular [3] and lung diseases [4], as well as lung carcinoma [5]. Nicotine induces oxidative stress primarily by stimulating the formation of reactive oxygen species (ROS). Furthermore, nicotine has detrimental effects on male reproductive health and fertility. Several studies demonstrated that nicotine disrupts spermatogenesis, reduces epididymal sperm count, decreases sperm motility, and compromises fertilizing capacity. Furthermore, this substance disrupts Leydig cell function, leading to decreased testosterone synthesis [6].
Accordingly, supplementation with substances possessing antioxidant and antiapoptotic properties is considered a promising therapeutic approach for the treatment of male infertility [7]. Phytotherapeutic agents and their bioactive compounds are attracting increasing attention as potential antioxidants [8]. Taraxasterol (3β,18α,18α)-Urs-20(30)-en-3-ol, a pentacyclic triterpene isolated from Taraxacum officinale (Asteraceae) [9], exhibits multiple protective effects, including inhibition of inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) expression in lipopolysaccharide-stimulated RAW264.7 macrophages and suppression of interleukin-1 beta (IL-1β)-induced nitric oxide (NO) and prostaglandin E2 (PGE2) production in human bladder cancer cells [10]. The antioxidant capacity of taraxasterol was confirmed in numerous studies. Expanding the use of traditional therapies and natural compounds for disease treatment requires reliable scientific verification of the corresponding mechanisms and efficacy [11]. Taraxasterol is a naturally occurring pentacyclic triterpenoid extracted primarily from T. officinale, a species widespread in temperate regions and long used in traditional herbal medicine for its wide range of purported therapeutic effects [11]. Historically, T. officinale has been used in traditional medicine to treat liver, digestive system, urinary tract, as well as inflammatory conditions, making it a valuable source of bioactive compounds, including taraxasterol. Pharmacological studies have since confirmed a number of these effects of taraxasterol, highlighting its potent anti-inflammatory, antioxidant, and anticancer properties, thereby making it a promising natural candidate for cytoprotective and therapeutic applications [9]. Including this information at the beginning of the study not only justifies the use of taraxasterol from a well-known plant source but also provides readers with insight into the rationale for its protective effects in models of oxidative stress-induced cellular toxicity, such as nicotine exposure in testicular cell lines. This structure highlights the translational potential of using a natural phytochemical derived from a widely available medicinal plant with established ethnopharmacological significance [12].
Consistent with the above findings, this study aimed to evaluate the protective effects of taraxasterol against nicotine-induced oxidative stress and apoptosis in murine testicular Leydig (TM3) and Sertoli (TM4) cell lines. Although previous studies have documented the antioxidant and cytoprotective effects of taraxasterol against toxic oxidative damage in various tissues, the associated protective role in testicular cells remains unexplored.
Material and methods
Materials used in the experiment
Murine testicular cell lines – TM3 (ATCC® CRL-1714™) and TM4 (ATCC® CRL-1715™) – were obtained from the Pasteur Institute (Tehran, Iran). Cells were cultured in Roswell Park Memorial Institute RPMI 1640 medium (Gibco, Cat. no. 11875-093) supplemented with 10% fetal bovine serum (FBS) (Gibco, Cat. no. 26140-079) and 1% penicillin-streptomycin solution (Gibco, Cat. no. 15140-122). Trypsin-EDTA (0.25%) for cell detachment was purchased from Sigma-Aldrich (Cat. no. T3924). MTT reagent used for the viability assay was acquired from Sigma-Aldrich (Cat. no. M2128) and prepared as a 5 mg/mL solution in phosphate-buffered saline (PBS) (Gibco, Cat. no. 10010023). Dimethyl sulfoxide (DMSO) for solubilizing formazan was procured from Merck (Cat. no. 100995). Nicotine hydrogen tartrate salt (for cytotoxicity experiments) was obtained from Sigma-Aldrich (Cat. no. N3876). Taraxasterol was attained from Sigma-Aldrich (Cat. no. SML0193). Malondialdehyde MDA assay kits were acquired from Cayman Chemical Company (Cat. no. 10009055). GSH levels were determined using reduced GSH standards (Sigma-Aldrich, Cat. no. G4251), glutathione S-transferase enzyme (GST) enzyme (Sigma-Aldrich, Cat. no. G9423), and buffer solutions from the GSH assay kit (Abcam, Cat. no. ab138881). ROS levels were measured using 2',7'-dichlorodihydrofluorescein diacetate (DCFH-DA) purchased from Sigma-Aldrich (Cat. no. D6883). Total RNA extraction kits were purchased from Qiagen (RNeasy Mini Kit, Cat. no. 74106), while reverse transcription kits were procured from Thermo Fisher Scientific (RevertAid First Strand cDNA Synthesis Kit, Cat. no. K1622). Quantitative real-time PCR was performed using SYBR Green Master Mix (Applied Biosystems, Cat. no. 4309155) with primers designed based on sequences from the National Center for Biotechnology Information (NCBI). Diphenylamine (DPA) reagent for DNA fragmentation analysis was prepared according to a standard protocol using chemical reagents obtained from Sigma-Aldrich (Cat. no. D8001). All materials and reagents were used according to the manufacturer’s instructions and laboratory quality standards to ensure the reliability and reproducibility of laboratory procedures [13].
TM3 and TM4 cell cultures
Murine testicular cell lines (TM3 for Leydig cells and TM4 for Sertoli cells) were cultured in RPMI 1640 medium supplemented with 10% FBS and 1% penicillin-streptomycin antibiotic solution. Cultures were maintained in sterile T-flasks under standard cell culture conditions at 37 °C in a humidified atmosphere containing 5% CO₂. Cells were subcultured at approximately 70-80% confluency to ensure optimal growth and viability. The medium was replaced every 2-3 days. During subculturing, cells were gently detached with 0.25% trypsin-EDTA in PBS and reseeded at the appropriate density to maintain logarithmic growth. All procedures were performed aseptically in a biological safety cabinet to prevent contamination. Cell morphology and viability were regularly monitored using phase-contrast microscopy to confirm epithelium-like traits typical of TM3 and TM4 cells [14].
Assessing the biological activity of taraxasterol
To quantify the cytotoxic or proliferative effects of taraxasterol on testicular cell viability, TM3 and TM4 cells were seeded at a density of 15,000 cells/well in sterile 96-well culture plates and allowed to adhere for 24 h under standard culture conditions (37 °C, 5% CO₂). The cells were then treated with serial dilutions of taraxasterol at concentrations of 0, 1, 2, 4, 8, 16, 32, and 64 μM prepared in complete culture medium and incubated for 24 h. Following treatment, 20 µL of MTT reagent (5 mg/mL in PBS) were added to each well, and the plates were incubated for 4 h at 37 °C to allow metabolically active cells to reduce MTT to insoluble formazan crystals. After incubation, the culture supernatant was carefully aspirated, and 100 µL of DMSO was added to each well to dissolve the formazan crystals. The plates were gently shaken for 10 minutes to ensure complete dissolution. Absorbance was then measured using a microplate spectrophotometer at 570-nm wavelength, using a reference wavelength of 630 nm to correct for background absorbance. Percent cell viability was calculated relative to untreated control cells. All assays were performed in triplicate to ensure reproducibility [15].
Nicotine cytotoxicity borderline (half-maximal inhibitory concentration, IC50)
Cells were cultured and maintained at 37 °C in a humidified incubator with 5% CO2. Before treatment, cells were seeded in 96-well plates at a density of 5,000 cells/well and allowed to attach and grow for 24 h. Nicotine stock solutions were prepared immediately before use and diluted in culture medium to achieve nine sequential concentrations: 0, 1, 2, 4, 8, 16, 32, 64, and 128 μM. After an initial 24-h incubation, the culture medium was replaced with medium containing the indicated nicotine concentrations. After treatment, cell viability was assessed using the MTT assay. Specifically, 20 μL of MTT reagent (5 mg/mL) was added to each well, followed by incubation at 37 °C for 4 h to form formazan crystals. The supernatant was carefully removed, and 150 µL of DMSO was added to each well to dissolve the formazan crystals. Absorbance was measured at 490 nm using a microplate reader. Absorbance values were normalized to the untreated control group (0 µM nicotine) to calculate the percentage of cell viability. Each concentration assay was performed in triplicate and repeated in three independent experiments. Dose-response curves were constructed by plotting nicotine concentration (logarithmic scale) against normalized cell viability. The IC50 value was calculated using nonlinear regression analysis [15].
Study groups and treatment protocol
The study comprised four groups, including a control group (cells cultured under standard conditions without exposure to taraxasterol or nicotine), a nicotine group (cells exposed to nicotine at doses of 81.67 μM and 62.64 μM for Leydig and Sertoli cells, respectively), a taraxasterol group (cells treated with taraxasterol at a dose of 32 μM), and a nicotine + taraxasterol group (cells pretreated with taraxasterol followed by exposure to nicotine to determine the compound’s potential protective effect against nicotine cytotoxicity).
Laboratory study design
To determine the potential protective effect of taraxasterol in mitigating nicotine-induced oxidative damage and apoptosis in testicular cell cultures, cells were seeded in 96-well plates and allowed to adhere to the substrate under standard culture conditions. The cells were then treated with taraxasterol (32 μM) for 24 h. Subsequently, the wells were exposed to nicotine at concentrations of 81.67 μM and 62.64 μM for Leydig and Sertoli cells, respectively. Finally, the cells were harvested, and several assays were performed to evaluate the potential protective effect of taraxasterol.
Cell viability assessment
The viability of the TM3 and TM4 cell lines was assessed using the MTT assay. Briefly, cells were seeded in 96-well plates at a density of 5-15×10³ cells/well and allowed to attach for 24 h under standard culture conditions (37 °C, 5% CO₂). After treatment with varying concentrations of nicotine and taraxasterol, 20 µL of MTT reagent (5 mg/mL in PBS) were added to each well, and the plates were incubated for 4 h at 37 °C to allow viable cells to metabolically reduce the yellow tetrazolium salt to insoluble purple formazan crystals. After incubation, the culture medium was carefully removed, and 100-150 µL of DMSO was added to each well to dissolve the formazan crystals. The plates were gently shaken for 10 min to ensure complete dissolution. Absorbance was measured at 570 nm using a reference wavelength of 630 nm by a microplate reader to quantify cell viability. Absorbance values were normalized relative to untreated control cells to calculate the percentage of viable cells [16].
MDA level measurement
Quantification of MDA, a biomarker of lipid peroxidation and oxidative stress, was performed using a commercially available MDA assay kit. Biological samples were prepared and placed in a 96-well microplate according to proper standards and controls. The assay is based on the reaction of MDA with thiobarbituric acid (TBA) to form a colored MDA-TBA adduct that exhibits a specific absorption peak. After sample incubation and reaction progression, absorbance was measured spectrophotometrically at 532 nm. MDA concentration was then calculated by interpolation from a standard curve constructed using known MDA concentrations. All samples and standards were analyzed in duplicate to ensure accuracy and reproducibility [17].
GSH level measurement
Cells were washed three times with PBS to remove residual culture medium and centrifuged at 1,500 rpm for 5 min. Cell pellets were lysed with 150 µLof 1X lysis buffer and vortexed. The lysates were incubated on ice for 15 min to facilitate protein solubilization. Samples were then centrifuged at 16,000 g (10 min, 4 °C) to pellet cellular debris, and the supernatant containing soluble intracellular components was collected for analysis. For GSH analysis, 5 µL of cell extract was dispensed into individual wells of a 96-well microplate, and 87.5 µL of buffer solution was added to each well. Standard curves were prepared in separate wells using reduced GSH standards at concentrations ranging from 0 to 10 nM. These standards were prepared by diluting a 1 mM GSH stock solution to final volumes of 1.3, 2.5, 5, and 10 µL/well, for a total volume of 92.5 µL. To initiate the enzymatic reaction, 5 µL of GST was added to all wells, including both samples and standards. Then, 2.5 µL of substrate solution was added, and the plate was incubated at 37 °C. Fluorescence intensity was measured using a microplate reader with excitation and emission wavelengths set to 360 nm and 485 nm, respectively. The concentration of reduced GSH in each sample was determined using a standard curve, with all measurements performed in duplicate to ensure reliability and accuracy [17].
Measurement of ROS-derived molecules
Cells were cultured in 96-well plates with black walls to minimize background fluorescence and allowed to adhere to the substrate before treatment. After the treatment period, the culture medium was removed, and the cells were gently washed with PBS to remove residual medium and unattached cells. Then, 100 µL of DCFH-DA diluted in fresh culture medium was added to the wells. The plates were incubated for 1 h at 37 °C in a humidified atmosphere to allow cellular uptake and intracellular deacetylation of DCFH-DA by esterase to the nonfluorescent compound DCFH. In the presence of intracellular ROS, DCFH was oxidized to the highly fluorescent dichlorofluorescein (DCF), which emitted fluorescence detectable at an excitation wavelength of 485 nm and an emission wavelength of 530 nm. Fluorescence intensity was measured using a fluorescence microplate reader. Fluorescence values obtained from untreated control wells were normalized to 1, and ROS levels in laboratory samples were expressed relative to the control [18].
Gene expression analysis
For RNA extraction, 1×10⁶ cells were placed in each well of 6-well plates. After treatment, cells were quickly harvested and immediately snap-frozen in liquid nitrogen to preserve RNA integrity. Total RNA was isolated using a commercial RNA isolation kit. RNA quality was assessed by agarose gel electrophoresis. The concentration and purity of RNA samples were determined spectrophotometrically by measuring the absorbance ratio at 260/280 nm and 230/260 nm using a NanoDrop spectrophotometer. Complementary DNA (cDNA) was synthesized from high-quality RNA using a reverse transcription kit according to the manufacturer’s instructions. The concentration and quality of the synthesized cDNA were verified using a NanoDrop spectrophotometer to ensure suitability for further applications. For quantitative PCR, the sequences of the target genes and the endogenous β-actin control (F: TACTGAGCTGCGTTTTACACC, R: TCCTGAGTCAAAAGCGCCAA) were retrieved from the NCBI database. Primers specific to these genes were designed using Gene Runner software and verified via NCBI BLAST to ensure specificity. Quantitative real-time PCR was performed using SYBR Green Master Mix in triplicate for each sample to ensure reproducibility. Relative quantification of gene expression levels (fold change) was performed using the comparative Ct method (2-ΔΔCt), normalizing SORT1 expression relative to glyceraldehyde-3-phosphate dehydrogenase. The genes were categorized into two groups of steroidogenic and antioxidant enzyme genes. The former included 3β-HSD (F: AGATAATCCTGAATGGCAACGA, R: TTTGCCCGTACAACCGAGA), 17β-HSD (F: TGGGTGCTGTGTTGGATGTGTGGCA, R: CAGTACACTTCGTGG), and StAR (F: TGCCTTAAATATCTCTAGCTC, R: TGCGTATCCTTCTGTGAGCC). The latter encompassed glutathione peroxidase 1 (GPx1) (F: ACAGTCCACCGTGTATGCC, R: CGTTCATCTCGGTGTAGTCCC), and the following superoxide dismutases (SOD): SOD1 (F: AAGCGGTGAACCAGTTGT, R: CCGGGCCACCAGTGTTCTTA), SOD2 (F: AGGAGAGTTGCTGGAGGCTA, R: TAGTAAGCGTGCTCCCACAC) and SOD3 (F: GAGAAGATAGGCGCAGCCGA, R: GAGAACCAAGCGCGTGATCT) [7].
Assessing DNA fragmentation rate (apoptosis)
Apoptosis was quantified by measuring the percentage of fragmented DNA using the DPA staining assay [19]. After laboratory treatments, 5×10⁶ cells were suspended in tris(hydroxymethyl)methylamino propanesulfonic acid-ethylenediaminetetraacetic acid (tris-EDTA-TAPS) buffer and centrifuged at 20,000 g for 10 min at 4 °C to separate intact chromatin (pellet) from fragmented DNA (supernatant). The supernatant containing fragmented DNA was carefully transferred to new microtubes (sample A). The pellet was resuspended in 1 mL of tris-EDTA-TAPS buffer and mixed with 1 mL of 25% trichloroacetic acid (TCA), then incubated overnight at 4 °C to precipitate DNA. After incubation, the samples were centrifuged again, and 160 μL of 5% TCA was added to each pellet to facilitate DNA hydrolysis. The samples were then heated at 90 °C for 15 minutes to cleave the DNA. After cooling, 320 μL of freshly prepared DPA reagent was added to each tube, and the samples were incubated at 37 °C for 4 h to develop a colorimetric reaction proportional to DNA fragmentation (sample B). Absorbance readings for samples A and B were measured at 600 nm using a spectrophotometer. The percentage of DNA fragmentation was calculated using the obtained absorbance values, providing a quantitative estimate of apoptotic DNA cleavage in the cell population, including DNA fragmentation (%) = [OD660(A)/OD660(B) − OD660(A)] × 100 [20].
Data analysis
Statistical analyses were conducted using SPSS software (v.19). One-way analysis of variance (ANOVA) was employed to compare differences among multiple groups, followed by Tukey’s post-hoc test for pairwise comparisons. Data are presented as mean ± standard deviation (SD). Statistical significance was determined at an appropriate p-value threshold (p<0.05), and all analyses met standard parametric assumptions [21].
Results
Bioactivity and IC50 assessment
The effects of varying concentrations of taraxasterol and nicotine on the viability of TM3 Leydig cells and TM4 Sertoli cells were assessed after 24-h exposure using the MTT assay. In Sertoli cells, taraxasterol treatment at concentrations up to 32 μM showed no significant effect on viability vs. the control, while a significant decrease was observed at 64 μM (89±3.12%, p<0.001), indicating potential cytotoxicity at high doses. Similarly, Leydig cells were not statistically significantly affected by taraxasterol at concentrations up to 32 μM, but viability was significantly reduced to 92±2.71% at 64 μM (p<0.001). Nicotine exposure caused a dose-dependent, significant decrease in cell viability in both cell lines: we observed a progressive reduction in Sertoli cell counts from 91±4.02% at 1 μM (p<0.005) to 36±1.92% at 128 μM (p<0.001), as well as in Leydig cells from 95±6.38% at 1 μM (p<0.05) to 40±2.28% at 128 μM (p<0.001). IC50 values were calculated as 81.67±5.34 μM for TM3 Leydig cells and 62.64±2.55 μM for TM4 Sertoli cells, indicating similar susceptibility of these cell types to nicotine cytotoxicity (Table 1).
Table 1. The effects of Taraxasterol (after 24 hours of exposure with various concentrations of 0-64 µM) and Nicotine (after 24 hours of exposure with various concentrations of 0-128 µM) on viability percentage (via MTT assay) of Sertoli (TM4) and Leydig (TM3) cell lines
|
|
Drugs |
Cell lines |
Taraxasterol or Nicotine concentration (µM) |
|||||||||
|
0 |
1 |
2 |
4 |
8 |
16 |
32 |
64 |
128 |
|
|||
|
Viability (%) |
Taraxasterol exposure |
Sertoli |
100±3.12 - |
100±1.18 NS |
100±2.40 NS |
100±3.22 NS |
100±4.02 NS |
100±1.98 NS |
100±0.89 NS |
89±3.12 ↓ p<0.001 |
NA |
|
|
Leydig |
100±4.10 - |
100±3.23 NS |
100±1.83 NS |
100±4.39 NS |
100±2.34 NS |
100±3.23 NS |
100±4.02 NS |
92±2.71 ↓ p<0.001 |
NA |
|
||
|
Nicotine exposure |
Sertoli |
100±2.54 - |
91±4.02 ↓ p<0.005 |
89±3.82 ↓ p<0.001 |
80±5.01 ↓ p<0.001 |
66±5.92 ↓ p<0.001 |
60±2.13 ↓ p<0.001 |
52±5.89 ↓ p<0.001 |
41±0.23 ↓ p<0.001 |
36±1.92 ↓ p<0.001 |
|
|
|
Leydig |
100±2.54 - |
95±6.38 ↓ p<0.05 |
90±3.35 ↓ p<0.05 |
83±4.51 ↓ p<0.001 |
78±2.91 ↓ p<0.001 |
72±5.32 ↓ p<0.001 |
65±4.33 ↓ p<0.001 |
51±2.01 ↓ p<0.001 |
40±2.28 ↓ p<0.001 |
|
||
Effect of taraxasterol on cell viability after nicotine exposure
Results of the cell viability study showed that nicotine exposure significantly reduced viability in both Leydig (TM3) and Sertoli (TM4) cells, with its values falling from 100% in the control group to 57% and 59%, respectively. In contrast, taraxasterol treatment increased cell viability to approximately 116% in TM3 cells and 120% in TM4 cells. Combined treatment with nicotine and taraxasterol restored cell viability to 87% in Leydig cells and 92% in Sertoli cells. These data demonstrate that taraxasterol exerts a potent cytoprotective effect, effectively compensating for the 30-35% loss of viability caused by nicotine toxicity (***p<0.001), and highlight its ability to maintain testicular cell survival under toxic conditions (Figure 1).
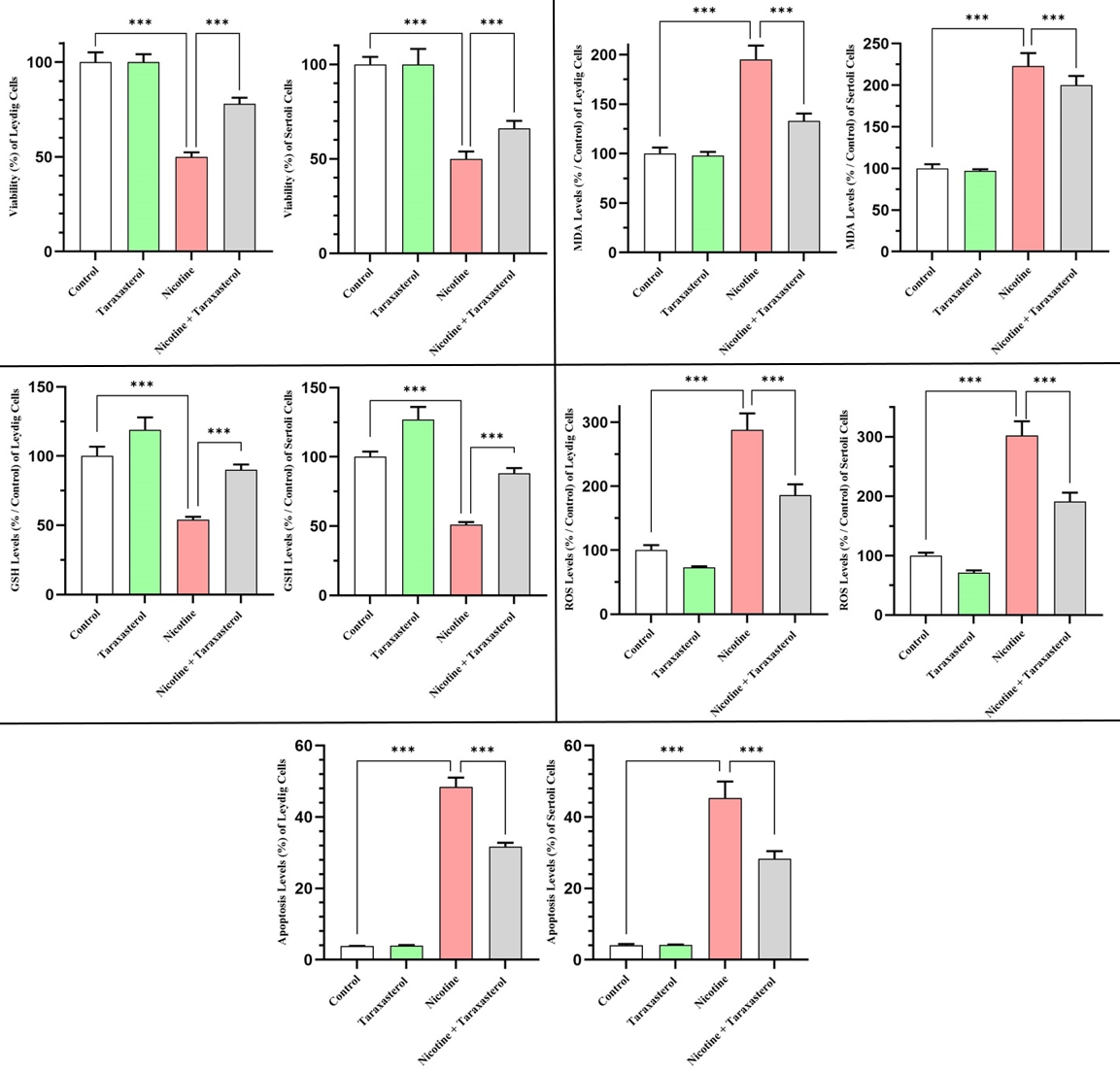
Figure 1. The effects of Taraxasterol on levels of cell viability, MDA, GSH, ROS, and apoptosis following cytotoxicity induced by nicotine exposure in both Leydig and Sertoli cell lines. All levels were represented as a percentage (%). *** indicates significant changes (p<0.001). White, green, red, and gray columns respectively refer to Control, Taraxasterol, Nicotine, and Nicotine + Taraxasterol groups. MDA: Malondialdehyde, GSH: Glutathione, and ROS: Reactive Oxygen Species.
Effect of taraxasterol on MDA levels after nicotine exposure
Oxidative stress analysis revealed that MDA levels, a marker of lipid peroxidation, significantly increased in TM3 and TM4 cells treated with nicotine (by 187% and 216%, respectively, relative to control). Taraxasterol alone had only a modest effect (110% of control for TM3 and 101% for TM4), while the nicotine + taraxasterol group restored MDA levels to 145% (Leydig cells) and 159% (Sertoli cells). Therefore, taraxasterol significantly reduced nicotine-induced lipid peroxidation by approximately 42% (Leydig cells) and 57% (Sertoli cells) vs. nicotine alone, thereby confirming its antioxidant capacity (Figure 1).
Effect of taraxasterol on GSH levels after nicotine exposure
Quantification of intracellular GSH levels revealed significant depletion after nicotine exposure: GSH levels decreased to 43% and 48% of control levels in Leydig and Sertoli cells, respectively. Taraxasterol increased GSH levels by 135% (Leydig cells) and 138% (Sertoli cells), while co-administration of nicotine restored GSH levels to values close to baseline (98% and 93%, respectively). These findings confirm that taraxasterol replenishes cellular antioxidant defenses and counteracts the oxidative stress caused by nicotine toxicity (Figure 1).
Effect of taraxasterol on ROS levels after nicotine exposure
ROS levels increased sharply under nicotine exposure, reaching 245% of control levels in Leydig cells and 294% in Sertoli cells. Taraxasterol alone reduced ROS levels by 86% (Leydig cells) and 80% (Sertoli cells), while the combination treatment group showed an intermediate reduction (173% in Leydig cells, 171% in Sertoli cells). This significant reduction indicates that taraxasterol significantly mitigates nicotine-induced oxidative stress and helps maintain cellular redox balance (Figure 1).
Effect of taraxasterol on apoptosis levels after nicotine exposure
Nicotine exposure sharply increased DNA fragmentation, which triggers apoptosis, reaching 37% (Leydig cells) and 26% (Sertoli cells) vs. a baseline level of approximately 4-5%. Taraxasterol-treated cells maintained low levels of apoptosis (5% in Leydig cells, 4% in Sertoli cells), while taraxasterol-treated cells showed a marked reduction in apoptosis (19% in Leydig cells, 12% in Sertoli cells). This confirms the antiapoptotic and protective effects of taraxasterol against nicotine-induced cell death in both testicular cell types (Figure 1).
Effect of taraxasterol on the expression of steroidogenic genes (3β-HSD, 17β-HSD, and StAR) after nicotine exposure
After 24 h of exposure, the relative expression of steroidogenic and antioxidant enzyme genes exhibited significant changes in both Leydig (TM3) and Sertoli (TM4) cells across the experimental groups. In the steroidogenic gene category, nicotine exposure significantly reduced the expression of 3β-HSD, 17β-HSD, and StAR in both cell types (p<0.05), reaching 0.67±0.02, 0.65±0.02, and 0.61±0.01 in TM3 cells and 0.66±0.04, 0.61±0.02, and 0.63±0.01 in TM4 cells vs. the control group. In contrast, treatment with taraxasterol alone significantly upregulated these genes, with expression levels increasing to 1.84±0.05, 1.67±0.06, and 2.03±0.06 in TM3 cells and to 1.56±0.03, 1.70±0.07, and 1.91±0.81 in TM4 cells (p<0.05). In cells pretreated with taraxasterol before nicotine exposure, a partial restoration of gene expression was observed, yielding intermediate values (1.33±0.03-1.46±0.04 in TM3 and 1.13±0.10-1.33±0.32 in TM4, p<0.05 vs. the nicotine-treated group), indicating a protective effect of taraxasterol against the suppression of steroidogenic pathways by nicotine. Analysis of antioxidant enzyme genes revealed a similar pattern. Nicotine caused a substantial downregulation of SOD1, SOD2, SOD3, and GPx1 in both Leydig and Sertoli cells (p<0.05), while taraxasterol treatment resulted in a marked increase in expression, especially in TM4 cells, where SOD1 and GPx1 showed an increase of 4.42±0.98 and 3.09±1.01 times, respectively. In the group receiving combined treatment with nicotine and taraxasterol, gene expression values were significantly higher than in the group receiving nicotine alone, indicating a reduction in oxidative stress. For instance, SOD1 and GPx1 levels in Leydig cells increased from 0.18±0.01 and 0.23±0.03 in the nicotine-treated group to 0.82±0.01 and 1.20±0.03 after taraxasterol pretreatment. Similarly, in Sertoli cells, SOD gene expression increased to 2.34±0.71-2.98±0.77 (p<0.05 vs. nicotine). Overall, these results indicate that nicotine significantly suppresses the expression of steroidogenic and antioxidant defense genes, leading to potential cytotoxic and oxidative effects in testicular cells, while taraxasterol administration not only enhances the baseline expression of these genes but also exerts significant cytoprotective and restorative effects against the nicotine-induced transcriptional decline (Table 2).
Table 2. Relative expression of steroidogenic and antioxidant enzyme genes in four groups of Control, Taraxasterol, Nicotine, and Nicotine + Taraxasterol
|
|
Genes |
Cells |
Study Groups |
|||
|
Control |
Taraxasterol |
Nicotine |
Nicotine + Taraxasterol |
|||
|
Steroidogenic Genes |
3β-HSD |
Leydig |
1 |
1.84±0.05 ƈ |
0.67±0.02 ƈ |
1.33±0.03 Ťŋ |
|
Sertoli |
1 |
1.56±0.03 ƈ |
0.66±0.04 ƈ |
1.13±0.10 Ťŋ |
||
|
17β-HSD |
Leydig |
1 |
1.67±0.06 ƈ |
0.65±0.02 ƈ |
1.35±0.04 Ťŋ |
|
|
Sertoli |
1 |
1.70±0.07 ƈ |
0.61±0.02 ƈ |
1.30±0.11 Ťŋ |
||
|
StAR |
Leydig |
1 |
2.03±0.06 ƈ |
0.61±0.01 ƈ |
1.46±0.04 Ťŋ |
|
|
Sertoli |
1 |
1.91±0.81 ƈ |
0.63±0.01 ƈ |
1.33±0.32 Ťŋ |
||
|
Antioxidant Enzyme Genes |
SOD1 |
Leydig |
1 |
2.32±0.45 ƈ |
0.18±0.01 ƈ |
0.82±0.01 Ťŋ |
|
Sertoli |
1 |
4.42±0.98 ƈ |
0.70±0.12 ƈ |
2.29±0.91 ƈŤŋ |
||
|
SOD2 |
Leydig |
1 |
1.67±0.03 ƈ |
0.04±0.01 ƈ |
1.45±0.04 Ťŋ |
|
|
Sertoli |
1 |
3.24±1.04 ƈ |
0.72±0.20 ƈ |
2.34±0.71 ƈŤŋ |
||
|
SOD3 |
Leydig |
1 |
3.49±1.81 ƈ |
0.78±0.02 ƈ |
1.03±0.33 Ťŋ |
|
|
Sertoli |
1 |
1.55±0.43 ƈ |
0.93±0.21 ƈ |
2.45±0.27 ƈŤŋ |
||
|
GPx1 |
Leydig |
1 |
2.79±0.15 ƈ |
0.23±0.03 ƈ |
1.20±0.03 Ťŋ |
|
|
Sertoli |
1 |
3.09±1.01 ƈ |
0.67±0.06 ƈ |
2.98±0.77 ƈŤŋ |
||
Effect of taraxasterol on the expression of antioxidant enzymes (SOD1, SOD2, SOD3, and GPx1) after nicotine exposure
Concerning antioxidant enzyme genes, nicotine significantly suppressed the expression of SOD1, SOD2, SOD3, and GPx1 in Leydig and Sertoli cells vs. the control group (p<0.05), consistent with impaired cellular defenses caused by oxidative stress. Conversely, taraxasterol alone significantly increased the transcription levels of these antioxidant enzymes (p<0.05), indicating enhanced antioxidant capacity. Furthermore, cells treated with nicotine and taraxasterol exhibited a significant increase in antioxidant gene expression, compared with nicotine-only treated cells (p<0.05), thereby suggesting that taraxasterol attenuates nicotine-induced oxidative damage by restoring antioxidant enzyme expression (Table 2).
Discussion
This study provided valuable insights into the protective effects of taraxasterol against nicotine cytotoxicity in Leydig (TM3) and Sertoli (TM4) cells, which are critical for maintaining male reproductive health. Given the widespread availability of nicotine and its documented detrimental effects on testicular function, identifying compounds that can mitigate this damage is essential for the development of potential therapeutic interventions. Our results demonstrated that taraxasterol plays a significant cytoprotective role by improving cell viability, reducing oxidative stress markers, and attenuating apoptosis in nicotine-exposed testicular cells. Specifically, taraxasterol alone was nontoxic to both cell types at low concentrations. In addition to maintaining cell viability, taraxasterol significantly reduced lipid peroxidation, as evidenced by decreased MDA levels, replenishment of intracellular GSH concentrations, and a reduction in intracellular ROS accumulation, thereby restoring the redox balance disrupted by nicotine. Importantly, taraxasterol also mitigated nicotine-induced apoptotic cell death, highlighting its potential to preserve testicular cell integrity. At the molecular level, key steroidogenic genes required for testosterone biosynthesis (including 3β-HSD, 17β-HSD, and StAR) were downregulated by nicotine and effectively upregulated by taraxasterol, indicating restoration of steroidogenic capacity. Similarly, the gene expression of antioxidant enzymes (SOD1, SOD2, SOD3, and GPx1) was impaired by nicotine but significantly restored by taraxasterol treatment, thereby highlighting its role in strengthening the antioxidant defense system.
In the present study, the cytotoxic effects of taraxasterol and nicotine were determined by the half-maximal IC50 using the MTT assay. Taraxasterol at concentrations up to 32 μM showed no significant toxicity in Sertoli and Leydig cells, maintaining cell viability at levels similar to control values, which was consistent with previously reported nontoxic doses of plant triterpenoids in similar testicular cell models [11]. However, at a concentration of 64 μM, taraxasterol caused a significant decrease in viability, indicating potential cytotoxicity at elevated concentrations, which was consistent with the known dose-dependent cytotoxicity thresholds observed for related phytochemicals. Nicotine exposure showed a clear dose-dependent cytotoxic effect on Leydig and Sertoli cells, highlighting the slightly higher sensitivity of Sertoli cells to nicotine toxicity. These results confirm previous studies illustrating the ability of nicotine to impair testicular cell viability through mechanisms involving oxidative stress and mitochondrial dysfunction [22]. Comparable IC50 values of Leydig and Sertoli cells suggest common pathways of susceptibility to nicotine cytotoxicity in the testicular microenvironment, thereby highlighting the key role of nicotine in compromising reproductive cell integrity. Mechanistically, nicotine’s toxic effects are associated with excessive ROS generation, depletion of endogenous antioxidants, and subsequent triggering of apoptotic cascades. The lack of cytotoxicity in taraxasterol at therapeutic concentrations confirms its suitability as a safe agent for modulating oxidative damage. Its cytoprotective potential is likely due to its ability to scavenge free radicals and enhance antioxidant defenses, as demonstrated in other models of oxidative damage [23]. Taken together, the IC50 assessment confirms the therapeutic range of taraxasterol and provides a basis for studying its protective mechanisms against nicotine-induced cellular damage in the male reproductive system.
Furthermore, both the control and taraxasterol-treated groups exhibited relatively high viability, indicating that taraxasterol alone does not negatively affect cell survival. In contrast, nicotine significantly reduced cell viability to approximately 50% in Leydig cells and 48% in Sertoli cells, illustrating its pronounced cytotoxic effects, consistent with previous reports linking nicotine toxicity to oxidative damage and mitochondrial dysfunction in testicular cells [24]. Notably, treatment with nicotine and taraxasterol significantly reduced cytotoxicity, restoring viability to approximately 80% in Leydig cells and 67% in Sertoli cells, confirming the protective role of taraxasterol in mitigating nicotine-induced damage. This finding is consistent with studies showing that natural triterpenoids can enhance cellular antioxidant defenses and inhibit apoptotic pathways, thereby suggesting that taraxasterol exerts its effects by reducing oxidative stress and maintaining mitochondrial integrity [25]. Mechanistically, taraxasterol’s antioxidant properties likely reduce ROS accumulation, stabilize cell membranes, and prevent apoptosis, thus maintaining cell viability despite nicotine exposure. These results support the results of previous studies demonstrating that phytochemicals can counteract the detrimental effects of nicotine on reproductive cells. For example, similar protective effects have been observed with compounds such as resveratrol and astaxanthin, which both mitigate oxidative stress and improve viability in nicotine-treated testicular models [26]. Differences in recovery rates between Leydig and Sertoli cells may reflect inherent differences in their susceptibility to oxidative damage and response to taraxasterol, requiring further investigation into their mechanisms of action. Taken together, our results highlight the potential of taraxasterol as a promising cytoprotective agent capable of preserving testicular cell function under nicotine exposure, providing insight into therapeutic strategies for nicotine-induced reproductive toxicity.
Lipid peroxidation is a critical indicator of oxidative damage in cells, with MDA serving as an established biomarker for this process. In the context of nicotine-induced cytotoxicity in Leydig (TM3) and Sertoli (TM4) cells, MDA levels were significantly elevated, indicating increased oxidative stress and cell membrane damage. Our study revealed that nicotine exposure resulted in an approximately twofold increase in MDA compared with the control and taraxasterol-only groups, confirming the potent prooxidant effects of nicotine. The latter was consistent with previous reports demonstrating the ability of nicotine to induce oxidative damage in testicular tissue. Notably, taraxasterol treatment significantly reduced MDA accumulation by approximately 30–35%, highlighting its antioxidant efficacy in mitigating lipid peroxidation. This reduction aligns with similar studies documenting the outstanding free radical scavenging properties of taraxasterol and its ability to preserve cell membrane integrity under oxidative stress [27]. The observed protective mechanism of taraxasterol likely involves modulation of endogenous antioxidant systems and direct suppression of ROS formation. By enhancing antioxidant enzyme activity and maintaining GSH levels, taraxasterol reduces the availability of substrates for lipid peroxidation, thereby inhibiting MDA formation. Such antioxidant effect is supported by data demonstrating a role for taraxasterol in activating the nuclear factor erythroid 2-related factor 2 (Nrf2)/antioxidant response element (ARE) pathway, which regulates cellular redox homeostasis and confers resistance to oxidative stress-induced apoptosis [28]. Collectively, these findings not only support previous studies of natural compounds that mitigate nicotine-induced reproductive toxicity but also highlight the therapeutic potential of taraxasterol in maintaining testicular cell integrity via attenuating oxidative stress-induced lipid damage.
Intracellular GSH plays a key role as the main antioxidant protecting cells from oxidative damage, and its depletion is closely associated with increased susceptibility to oxidative stress and cellular damage [29]. In this study, nicotine exposure caused a marked and statistically significant decrease in GSH levels in both Leydig and Sertoli cells to approximately 50% of control values, highlighting the detrimental effects of nicotine on cellular redox balance. These results are consistent with previous reports demonstrating that nicotine induces oxidative stress by excessively consuming cellular antioxidants, particularly GSH, thereby impairing cellular defense mechanisms [30]. In contrast, treatment with taraxasterol alone increased GSH concentrations above baseline values, indicating its intrinsic ability to enhance antioxidant status in testicular cells. This finding is consistent with previous studies highlighting the antioxidant potential of taraxasterol and related pentacyclic triterpenes in enhancing the activity of endogenous antioxidant systems, including the GSH synthesis pathway [31]. Importantly, simultaneous administration of taraxasterol and nicotine resulted in a significant restoration of GSH levels to nearly 90% of control concentrations, demonstrating its ability to replenish antioxidant stores depleted by nicotine-induced oxidative damage. The underlying mechanisms of this protective effect are likely related to taraxasterol’s ability to modulate key antioxidant regulatory pathways, such as activation of the Nrf2 signaling cascade, which mediates transcriptional activation of GSH biosynthetic and recycling enzymes [32]. Moreover, taraxasterol may exert an indirect antioxidant effect by attenuating ROS formation, thereby reducing GSH consumption and maintaining redox homeostasis. These mechanistic findings are supported by parallel studies of similar phytochemicals, which have been shown to counteract toxin-induced oxidative stress through GSH modulation in reproductive tissues [33].
Elevated intracellular ROS levels represent a central mechanism underlying the cytotoxic effects of nicotine on testicular cells, contributing to oxidative stress-mediated cell damage. Our results showed that nicotine exposure led to a nearly threefold increase in ROS production in both Leydig (TM3) and Sertoli (TM4) cell lines, highlighting the oxidative stress imposed by nicotine. Notably, treatment with taraxasterol alone significantly reduced ROS levels vs. the control group, which is consistent with its established antioxidant properties reported in previous phytochemical studies [34]. This strong inhibition of ROS formation suggests that taraxasterol actively neutralizes free radicals or enhances endogenous antioxidant defenses. Importantly, treatment with nicotine and taraxasterol significantly suppressed ROS accumulation by approximately 30-40% vs. cells treated with nicotine alone, indicating a substantial reduction in nicotine-induced oxidative stress. These observations support similar reports in which taraxasterol and related pentacyclic triterpenoids modulated oxidative stress biomarkers by inducing activation of the Nrf2 pathway and increasing the activity of antioxidant enzymes such as SOD and GSH peroxidase. By restoring redox homeostasis, taraxasterol likely prevents the subsequent pathological consequences of excess ROS, including lipid peroxidation, DNA damage, and apoptosis in reproductive tissues. Taken together, these data support the potential of taraxasterol as a natural antioxidant capable of protecting testicular cells from nicotine-induced oxidative damage, both through direct free radical scavenging and by enhancing cellular antioxidant systems [35].
This study clearly demonstrated the effect of taraxasterol on apoptosis after the cytotoxic effect of nicotine, providing important insights into its cytoprotective mechanism in Leydig (TM3) and Sertoli (TM4) cells. Quantitative DNA fragmentation analysis revealed negligible apoptosis in the control and taraxasterol-only groups, confirming minimal basal cell death in the absence of toxic exposure. However, nicotine exposure causes a significant increase in apoptosis, with its level reaching nearly 50% in both cell types, reflecting the known proapoptotic and cytotoxic effects of nicotine, which have been consistently observed in previous studies. The significant reduction in apoptosis to approximately 30% after combination treatment with nicotine and taraxasterol highlights the potent antiapoptotic capacity of taraxasterol, which is consistent with previous findings where triterpenoids exerted protective effects by modulating cell death pathways. This protective effect is likely mediated by the inhibition of mitochondria-dependent apoptotic signaling, including reduced caspase activation and preservation of mitochondrial membrane potential, as supported by mechanistic studies of taraxasterol and structurally related compounds. Furthermore, the antioxidant properties of taraxasterol contribute to its antiapoptotic effects by reducing oxidative stress, the latter being a major trigger of apoptosis in nicotine-induced testicular toxicity [36]. By significantly reducing ROS accumulation and lipid peroxidation, as demonstrated by decreased intracellular ROS and MDA levels, taraxasterol restores redox homeostasis, thereby preventing oxidative DNA damage that leads to programmed cell death. Taken together, these data suggest that taraxasterol’s dual ability to stabilize mitochondrial function and enhance the cellular antioxidant defense system provides a potent cytoprotective effect against nicotine-induced apoptosis. This study adds to the growing evidence that taraxasterol is a promising candidate for intervention in nicotine-induced male reproductive toxicity and associated oxidative damage.
This study highlights a critical aspect of nicotine-induced reproductive toxicity, viz., its detrimental effects on key steroidogenic genes in testicular Leydig and Sertoli cells, which are essential for steroid hormone biosynthesis. Consistent with previous studies demonstrating the ability of nicotine to impair steroidogenesis by downregulating enzymes such as 3β-HSD, 17β-HSD, and StAR, our findings demonstrate significant suppression of these gene expression upon nicotine exposure in both cell lines. This suppression likely contributes to impaired testosterone synthesis and testicular dysfunction, confirming the findings of Nabi et al. (2017), who reported similar reductions in steroidogenic enzymes in nicotine toxicity [37]. Importantly, taraxasterol treatment significantly upregulated the expression of these genes vs. their control levels, suggesting not only a reversal of the inhibitory effect of nicotine but also a potential stimulatory role in steroidogenesis. This is consistent with previous studies on triterpenoids, where taraxasterol-related compounds have been shown to promote steroidogenic activity through modulation of intracellular signaling pathways associated with hormone synthesis [38]. The protective effects of taraxasterol against nicotine-induced steroidogenesis impairment were further confirmed by the partial restoration of the expression of these enzymes in treated cells. Mechanistically, the anti-inflammatory and antioxidant properties of taraxasterol may attenuate oxidative stress and cytokine-mediated transcriptional suppression of steroidogenic genes. By reducing ROS accumulation and inflammatory signaling cascades triggered by nicotine, taraxasterol likely preserves the transcriptional machinery critical for steroidogenesis. This hypothesis is supported by additional data from studies of natural compounds such as resveratrol and curcumin, which exert similar protective effects on steroidogenic genes through restoration of redox balance and inhibition of the NF-κB pathway [39].
The present study demonstrated that nicotine exposure significantly suppresses the expression of key antioxidant enzyme genes, including SOD1, SOD2, SOD3, and GPx1, in both Leydig (TM3) and Sertoli (TM4) cells vs. untreated controls, reflecting impaired cellular antioxidant defenses induced by oxidative stress. This observation is closely consistent with previous studies indicating that nicotine promotes oxidative damage by suppressing the activity of critical antioxidant enzymes, thereby exacerbating ROS accumulation and cellular dysfunction [40]. In contrast, taraxasterol treatment alone significantly upregulates the transcriptional levels of these antioxidant genes, consistent with its known antioxidant and cytoprotective properties observed in natural compounds with phytochemical activity [41]. Importantly, cells treated with nicotine and taraxasterol demonstrated a significant restoration of antioxidant gene expression compared to nicotine treatment alone, indicating that taraxasterol effectively mitigates nicotine-induced oxidative stress by restoring antioxidant defense mechanisms. Mechanistically, increased expression of SOD1, SOD2, and SOD3 (superoxide dismutases responsible for catalyzing the dismutation of superoxide radicals into hydrogen peroxide), coupled with increased expression of GPx1, which promotes hydrogen peroxide reduction, reflects a comprehensive improvement in cellular redox homeostasis. Taraxasterol’s bioactive compounds likely activate the Nrf2-ARE signaling pathway, which regulates the expression of these antioxidant enzymes, thereby enhancing the cell’s ability to detoxify ROS and prevent oxidative damage. These data support existing literature that natural phytochemicals exert protective effects against environmental toxicants through modulation of oxidative signaling pathways [42].
Our study highlighted the central role of the Nrf2 signaling pathway in mediating the cytoprotective effects of taraxasterol. The marked activation of antioxidant genes (particularly SOD1 and GPx1) in taraxasterol-treated groups, even in the absence of nicotine, strongly suggests that activation of the Nrf2 pathway is a fundamental mechanism underlying the protective effect of taraxasterol [11]. This transcription factor, when activated, dissociates from its cytoplasmic inhibitor Kelch-like ECH-associated protein 1 (Keap1), translocates to the nucleus, and binds to AREs in the promoter regions of target genes, thereby inducing a potent antioxidant response, including enhanced synthesis of phase II detoxification enzymes and key antioxidant molecules such as heme oxygenase-1 (HO-1) and GPx family members. Our experimental data are consistent with this paradigm. Increased basal expression of SOD1 and GPx1 reflects a taraxasterol-enhanced antioxidant environment that prepares cells by significantly reducing ROS accumulation and subsequent oxidative damage, as evidenced by lower MDA levels and preservation of intracellular glutathione (GSH) [31]. The biological consequences of this redox homeostasis are multifaceted: reduced oxidative stress attenuates apoptosis (as evidenced by reduced DNA fragmentation) and promotes the restoration of expression of steroidogenic genes critical for Leydig and Sertoli cell function. These mechanistic data are strongly consistent with previous pharmacological studies demonstrating taraxasterol-mediated activation of the Nrf2 pathway in various experimental models, including neuronal, hepatic, and cardiac tissues, where taraxasterol promotes nuclear Nrf2 accumulation and transcriptional activation of antioxidant genes to promote cytoprotection and limit inflammatory responses. Therefore, it is plausible, even compelling, to hypothesize that the primary mechanism of taraxasterol’s action in mitigating nicotine toxicity is through activation of the Nrf2 pathway, which orchestrates a coordinated program of cellular defense. This unified mechanism elegantly explains the parallel upregulation of antioxidant gene expression, reduction in ROS and lipid peroxidation, GSH preservation, suppression of apoptosis, and normalization of steroidogenesis observed in our Leydig and Sertoli cell models. Integration of this hypothesis into the discussion provides a compelling conceptual framework linking molecular, biochemical, and functional data and significantly strengthens the scientific narrative [43].
Limitations of the Study and Implications for Future Research
Despite the significant results demonstrating the cytoprotective effect of taraxasterol against nicotine toxicity in Leydig (TM3) and Sertoli (TM4) cells, several limitations of this study should be noted. First, the study was conducted exclusively in vitro, limiting the direct extrapolation of the results to in vivo systems where complex physiological interactions occur. The systemic toxicity and metabolism of nicotine, as well as the bioavailability and pharmacokinetics of taraxasterol, may differ significantly in animal models or in humans compared to cell culture conditions. Furthermore, this study focused on two testicular cell lines representing Leydig and Sertoli cells and did not examine the effects on germ cells or the broader testicular microenvironment, which are critical for healthy reproductive function.
Future studies should expand molecular investigations, including proteomic and transcriptomic profiling, to elucidate the detailed pathways involved in taraxasterol-mediated cytoprotection.
Conclusion
This study demonstrated that taraxasterol exerts significant cytoprotective effects against nicotine toxicity in Leydig (TM3) and Sertoli (TM4) cell lines. Importantly, taraxasterol treatment significantly attenuates the cytotoxic effects of nicotine, increases cell viability, and restores redox balance, as evidenced by decreased MDA and ROS levels, along with replenished GSH content. Furthermore, taraxasterol mitigates nicotine-induced apoptosis and reverses the downregulation of key steroidogenic and antioxidant enzyme genes, highlighting its multifaceted protective mechanisms. Overall, these results support the potential of taraxasterol as a therapeutic agent to mitigate nicotine-associated testicular toxicity by enhancing cellular antioxidant defense and preserving steroidogenic function, which requires further in vivo and clinical studies.
Abbreviations
MDA, malondialdehyde; GSH, glutathione; ROS, reactive oxygen species; iNOS, inducible nitric oxide synthase; COX-2, cyclooxygenase-2; IL-1β, interleukin-1 beta; NO, nitric oxide; PGE2, prostaglandin E2; RPMI, Roswell Park Memorial Institute; FBS, fetal bovine serum; PBS, phosphate-buffered saline; DMSO, dimethyl sulfoxide; DCFH-DA, 2’;7’-dichlorodihydrofluorescein diacetate; TBA, thiobarbituric acid; GST, glutathione S-transferase; DCF, 2’,’-dichlorofluorescein; cDNA, complementary DNA; DPA, diphenylamine; tris-EDTA-TAPS, tris(hydroxymethyl)methylamino propanesulfonic acid-ethylenediaminetetraacetic acid; TCA, trichloroacetic acid; SD, standard deviation.
Author contributions
Conceptualization: FKhHA; methodology: IR; software: AGh; data processing: FKhHA; investigation: MHP; validation: FKhHA; formal analysis: IR; supervision: FKhHA; funding acquisition: FKhHA; visualization: FKhHA; project administration: IR; resources: MSR; writing: MHL.
Funding
Financial support was provided by the Research Council of Kermanshah University of Medical Sciences (No. 4030687).
Availability of data and materials
The datasets used and analyzed in this study are available from the corresponding author upon reasonable request.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Conflict of interest
The authors declare that they have no conflicts of interest.
- Eisenberg ML, Esteves SC, Lamb DJ, Hotaling JM, Giwercman A, Hwang K, et al. Male infertility. Nat Rev Dis Prim 2023; 9(1): 49. https://doi.org/10.1038/s41572-023-00459-w.
- Hemmatabadi FK, Partovyan M, Chavoshinezhad N, Keivan M, Dadfar R, Maryamneghari SM, et al. Immediate intratesticular injection of circulating blood serum-derived exosomes may alleviate orchitis caused by cisplatin. Rep Develop Med 2025; 9(2): 75-82. https://doi.org/10.1097/RD9.0000000000000103.
- Salari N, Rahimi S, Darvishi N, Abdolmaleki A, Mohammadi M. The global prevalence of E-cigarettes in youth: A comprehensive systematic review and meta-analysis. Public Health Pract (Oxf) 2024; 7: 100506. https://doi.org/10.1016/j.puhip.2024.100506.
- Mo R, Zhang J, Chen Y, Ding Y. Nicotine promotes chronic obstructive pulmonary disease via inducing pyroptosis activation in bronchial epithelial cells. Mol Med Rep 2022; 25(3): 92. https://doi.org/10.3892/mmr.2022.12608.
- Leiter A, Veluswamy RR, Wisnivesky JP. The global burden of lung cancer: current status and future trends. Nat Rev Clin Oncol 2023; 20(9): 624-639. https://doi.org/10.1038/s41571-023-00798-3.
- Barbagallo F, Assenza MR, Torrisi F, Buonacquisto A, Pallotti F. The smoky impact of nicotinic acetylcholine receptors on testicular function. J Clin Med 2024; 13(17): 5097. https://doi.org/10.3390/jcm13175097.
- Jalili C, Khani F, Salahshoor MR, Roshankhah S. Protective effect of curcumin against nicotine-induced damage on reproductive parameters in male mice. Int J Morphol 2014; 32(3): 844-849. https://doi.org/10.4067/S0717-95022014000300017.
- Salahshoor MR, Abdolmaleki A, Faramarzi A, Jalili C, Shiva R. Does Tribulus terrestris improve toxic effect of malathion on male reproductive parameters? J Pharm Bioallied Sci 2020; 12(2): 183-191. https://doi.org/10.4103/jpbs.JPBS_224_19.
- Obafemi OT, Ayeleso AO, Obafemi BA, Adewale OB, Omiyale BO, Lebelo SL, et al. Pharmacological relevance of taraxasterol: A review. Pharmacological Research – Modern Chinese Medicine 2024; 13: 100533. https://doi.org/10.1016/j.prmcm.2024.100533.
- Wang Q, Zhang R, He Y, Mao G, Kong Z. Taraxasterol enhanced bladder cancer cells radiosensitivity via inhibiting the COX-2/PGE2/JAK2/STAT3/MMP pathway. Int J Radiat Biol 2024; 100(5): 791-801. https://doi.org/10.1080/09553002.2024.2324475.
- Jiao F, Tan Z, Yu Z, Zhou B, Meng L, Shi X. The phytochemical and pharmacological profile of taraxasterol. Front Pharmacol 2022; 13: 927365. https://doi.org/10.3389/fphar.2022.927365.
- Wu J, Sun J, Liu M, Zhang X, Kong L, Ma L, et al. Botany, traditional use, phytochemistry, pharmacology and quality control of Taraxaci herba: Comprehensive review. Pharmaceuticals (Basel) 2024; 17(9): 1113. https://doi.org/10.3390/ph17091113.
- Ahmadi F, Pazhouhi M, Bakhtiari M, Khani-Hematabadi F, Ghanbari A, Gholami M, et al. The effect of harmine on dental pulp stem cells differentiation into neural cells in two-dimensional and three-dimensional cell cultures. Jundishapur J Nat Pharm Prod 2023; 18(4): e135563. https://doi.org/10.5812/jjnpp-135563.
- Jalili C, Hemmatabadi FK, Bakhtiyari M, Abdolmaleki A, Moradi F. Effects of three-dimensional sodium alginate scaffold on maturation and developmental gene expressions in fresh and vitrified preantral follicles of mice. Int J Fertil Steril 2021; 15(3): 167-177. https://doi.org/10.22074/ijfs.2020.134609.
- Srinivasan B, Lloyd MD. Dose-Response Curves and the Determination of IC50 and EC50 Values. J Med Chem 2024; 67(20): 17931-17934. https://doi.org/10.1021/acs.jmedchem.4c02052.
- Buranaamnuay K. The MTT assay application to measure the viability of spermatozoa: A variety of the assay protocols. Open Vet J 2021; 11(2): 251-269. https://doi.org/10.5455/OVJ.2021.v11.i2.9.
- Zhang M, Ma B, Yang S, Wang J, Chen J. Bisphenol A (BPA) induces apoptosis of mouse Leydig cells via oxidative stress. Environ Toxicol 2023; 38(2): 312-321. https://doi.org/10.1002/tox.23690.
- Jambor T, Zajickova T, Arvay J, Ivanisova E, Tirdilova I, Knizatova N, et al. Exceptional properties of Lepidium sativum L. extract and its impact on cell viability, ROS production, steroidogenesis, and intracellular communication in mice Leydig cells in vitro. Molecules 2022; 27(16): 5127. https://doi.org/10.3390/molecules27165127.
- Ghanbari A, Jalili C, Abdolmaleki A, Shokri V. Effects of cisplatin and acacetin on total antioxidant status, apoptosis and expression of OCTN3 in mouse testis. Biotech Histochem 2022; 97(3): 185-191. https://doi.org/10.1080/10520295.2021.1925347.
- Abdolmaleki A, Pazhouhi M, Rashidi I, Jalili C, Heshmati S, Khani-Hemmatabadi F. Molecular Basis of Apoptosis Induced by Taraxasterol on Human Melanoma Cell Line Growth Inhibition: An in-Vitro Study. J Adv Med Biom Res 2024; 32(153): 288-898. http://journal.zums.ac.ir/article-1-7415-fa.html.
- Roshankhah S, Abdolmaleki A, Salahshoor MR. Anti-inflammatory, antiapoptotic, and antioxidant actions of Middle Eastern Phoenix dactylifera extract on mercury-induced hepatotoxicity in vivo. Mol Biol Rep 2020; 47(8): 6053-6065. https://doi.org/10.1007/s11033-020-05680-4.
- East N, Bishop E, Breheny D, Gaca M, Thorne D. A screening approach for the evaluation of tobacco-free ‘modern oral’ nicotine products using Real Time Cell Analysis. Toxicol Rep 2021; 8: 481-488. https://doi.org/10.1016/j.toxrep.2021.02.014.
- Zhang Y, Shaari RB, Awang Nawi MAB, Hassan AB, Cui C. Pharmacological action and research progress of taraxasterol. Curr Pharm Biotechnol 2024; 25(14): 1767-1777. https://doi.org/10.2174/0113892010276692231220103636.
- Yuan Y, Li Z, Li M, Jin T, Zhang X, Liu X, et al. Mitochondria oxidative stress mediated nicotine-promoted activation of pancreatic stellate cells by regulating mitochondrial dynamics. Toxicol In Vitro 2022; 84: 105436. https://doi.org/10.1016/j.tiv.2022.105436.
- Asgari R, Mansouri K, Abdolmaleki A, Bakhtiari M. Association of matrix metalloproteinases with male reproductive functions; with focus on MMP2, 7, and 9. Meta Gene 2021; 29: 100906. https://doi.org/10.1016/j.mgene.2021.100906.
- Marzagalli M, Battaglia S, Raimondi M, Fontana F, Cozzi M, Ranieri FR, et al. Anti‐inflammatory and antioxidant properties of a new mixture of vitamin C, collagen peptides, resveratrol, and astaxanthin in tenocytes: Molecular basis for future applications in tendinopathies. Mediators Inflamm 2024; 2024: 5273198. https://doi.org/10.1155/2024/5273198.
- Yan KX, Ge BJ, Sang R, Zhao P, Liu XM, Yu MH, et al. Taraxasterol attenuates zearalenone-induced kidney damage in mice by modulating oxidative stress and endoplasmic reticulum stress. Ecotoxicol Environ Saf 2024; 285: 117093. https://doi.org/10.1016/j.ecoenv.2024.117093.
- Li Z, Lian Y, Wei R, Jin L, Cao H, Zhao T, et al. Effects of taraxasterol against ethanol and high-fat diet-induced liver injury by regulating TLR4/MyD88/NF-κB and Nrf2/HO-1 signaling pathways. Life Sci 2020; 262: 118546. https://doi.org/10.1016/j.lfs.2020.118546.
- Roshankhah S, Abdolmaleki A, Jalili C, Salahshoor MR. Crocin reduces oxidative stress produced by busulfan administration on fertility of male rats. J Adv Med Biomed Res 2019; 27(125): 25-33. https://doi.org/10.30699/jambs.27.125.25.
- Nimbal A, Ahirrao B, Vishwakarma A, Vishwakarma P, Wani AB, Patil AA. Comparative evaluation of GSH, total protein and albumin levels in patients using smokeless tobacco with oral precancerous and cancerous lesions. Med Int (Lond) 2024; 4(2): 15. https://doi.org/10.3892/mi.2024.139.
- Xu L, Yu Y, Sang R, Li J, Ge B, Zhang X. Protective effects of taraxasterol against ethanol‐induced liver injury by regulating CYP2E1/Nrf2/HO‐1 and NF‐κB signaling pathways in mice. Oxid Med Cell Longev 2018; 2018: 8284107. https://doi.org/10.1155/2018/8284107.
- Salahshoor MR, Abdolmaleki A, Faramarzi A, Ziapour A, Roshankhah S. Thymus vulgaris attenuates Myleran-induced reproductive damage by decreasing oxidative stress and lipid peroxidation in male rats. J Hum Reprod Sci 2020; 13(1): 38-45. https://doi.org/10.4103/jhrs.JHRS_134_19.
- Jalili C, Abdolmaleki A, Roshankhah S, Salahshoor MR. Effects of gallic acid on rat testopathy following morphine administration: An experimental study. J Herbmed Pharmacol 2020; 9(1): 61-67. https://doi.org/10.15171/jhp.2020.09.
- Xueshibojie L, Duo Y, Tiejun W. Taraxasterol inhibits cigarette smoke-induced lung inflammation by inhibiting reactive oxygen species-induced TLR4 trafficking to lipid rafts. Eur J Pharmacol 2016; 789: 301-307. https://doi.org/10.1016/j.ejphar.2016.07.047.
- Almagro L, Calderón AA, Pedreño MA, Ferrer MA. Differential response of phenol metabolism associated with antioxidative network in elicited grapevine suspension cultured cells under saline conditions. Antioxidants (Basel) 2022; 11(2): 388. https://doi.org/10.3390/antiox11020388.
- Meng TT, Wang W, Meng FL, Wang SY, Wu HH, Chen JM, et al. Nicotine causes mitochondrial dynamics imbalance and apoptosis through ROS mediated mitophagy impairment in cardiomyocytes. Front Physiol 2021; 12: 650055. https://doi.org/10.3389/fphys.2021.650055.
- Nabi M, Rasool SUA, Ashraf S, Andrabi SM, Amin S. Androgen receptor signalling transactivator lncRNAs PRNCR1 and PCGEM contribute to PCOS pathogenesis. 2023; PREPRINT (Version 1). https://doi.org/10.21203/rs.3.rs-2598360/v1.
- Mohammadi-Cheraghabadi M, Hazrati S. Terpenoids, steroids, and phenolic compounds of medicinal plants. In: Arora C, Verma DK, Aslam J, Mahish PK, Eds. Phytochemicals in Medicinal Plants: Biodiversity, Bioactivity and Drug Discovery. Berlin, Boston: De Gruyter. 2023: 105-130. https://doi.org/10.1515/9783110791891-005.
- Ferlazzo N, Micali A, Marini HR, Freni J, Santoro G, Puzzolo D, et al. A flavonoid-rich extract from bergamot juice, alone or in association with curcumin and resveratrol, shows protective effects in a murine model of cadmium-induced testicular injury. Pharmaceuticals (Basel) 2021; 14(5): 386. https://doi.org/10.3390/ph14050386.
- Lewandowski Ł, Kepinska M, Milnerowicz H. Alterations in concentration/activity of superoxide dismutases in context of obesity and selected single nucleotide polymorphisms in genes: SOD1, SOD2, SOD3. Int J Mol Sci 2020; 21(14): 5069. https://doi.org/10.3390/ijms21145069.
- Li H, Sang R, Zhao X, Li C, Wang W, Wang M, et al. Research note: Taraxasterol alleviates aflatoxin B1-induced oxidative stress in chicken primary hepatocytes. Poult Sci 2023; 102(1): 102286. https://doi.org/10.1016/j.psj.2022.102286.
- Ge B, Sang R, Wang W, Yan K, Yu Y, Kong L, et al. Protection of taraxasterol against acetaminophen-induced liver injury elucidated through network pharmacology and in vitro and in vivo experiments. Phytomedicine 2023; 116: 154872. https://doi.org/10.1016/j.phymed.2023.154872.
- Zhang X, Xiong H, Liu L. Effects of taraxasterol on inflammatory responses in lipopolysaccharide-induced RAW 264.7 macrophages. J Ethnopharmacol 2012; 141(1): 206-211. https://doi.org/10.1016/j.jep.2012.02.020.
Received 6 September 2025, Revised 12 November 2025, Accepted 27 November 2025
© 2025, Russian Open Medical Journal
Correspondence to Fuzieh Khani Hemmatabadi. E-mail: fuziekhani@yahoo.com.